Conflicts Between Darwin and Geological Succession
Prior to the time The Origin of Species was written, the geologic time
scale in its modern form was already fully developed. In fact, the time
scale based on fossils was built by geologists who were creationists (Raup,
1981). Scientists of Darwin's day did not equate geologic succession with
evolution, nor should we today. One reason is that geological succession
does not reveal how new species came into existence, it only reveals when.
Another reason is that the order of appearance found in the fossil record
is, as a general rule, systematically backwards from the major predictions
of Darwinian theory.
Geological succession is often looked upon as primary evidence for the fact
of evolution. Amoebas appear before fish, and fish before philosophers.
In the popular view, this succession from simple to complex is evidence
for evolution. Geological succession, however, does not necessarily proceed
from simple to complex. For example, trilobites, among the most advanced
of the arthropods, are the first arthropods to appear in the geological
record. Darwin lamented over the complexity of the vertebrate eye, confessing
that "to suppose that the eye...could have been formed by natural
selection, seems, I freely confess, absurd in the highest possible degree."
(Darwin, 1859, p. 217) Yet some scientists believe that the schizochroal
eyes of some trilobites possessed the most sophisticated optical systems
ever utilized by any organism (Levi-Setti, 1975). David Raup has stated
that the trilobites" ... used an optical design which would require
a well trained and imaginative optical engineer to develop today ... " (Raup,
1979). The early metazoans were anything but simple.
The complexity of the early metazoans, however, is but a wrinkle in the
irony of Darwinism. The real irony lies in the general systematic order
of appearance of the taxa in the fossil record.
Life is far too complex for our theories to predict the specific body plans
which should come into existence as the result of evolution. Both Darwinian
gradualism and punctuated equilibrium, however, predict that a systematic
branching pattern should be evident if all life has arisen from a common
ancestor (see Figures 2 and 3). Both of these models are based upon relatively
small intra- or inter-species change and, therefore, predict that numerous
transitional species must come into existence which gradually diverge to
produce the disparity of the higher taxa. Both models predict that as new
species emerge and morphological distance between them increases, new genera
and eventually new families should appear. As increasing diversity occurs,
new orders, new classes, and ultimately new phyla should come into existence.
In short, diversity should precede disparity. The gradually increasing diversity
of the lower taxa should ultimately result in the disparity of the higher
taxa and the appearance of major new body plans. The pattern of geological
succession predicted by both of these evolutionary theories is from bottom
to top: species to genera to families to orders to classes to phyla.

FIGURE 2: Phyletic gradualism
Through the continuously gradual transformation of species this model predicts
that the increasing diversity of the lower taxa should precede the disparity
of the higher taxa.

FIGURE 3: Punctuated Equilibrium
Under this model evolutionary change is confined to shorter time spans and
small isolated populations in order to account for the lack of transitional
forms among the lower taxa. Lower taxon-level punctuations, however, require
numerous transitional steps to produce the disparity of the higher taxa.
This model also predicts that the increasing diversity of the lower taxa
should precede the disparity of the higher taxa.
The pervasive pattern of geological succession is contrary to these
theories (see Figure 4). Disparity precedes diversity. The initial appearance
of virtually all phyla occurs with very low species diversity. The origin
of the major body plans is not the result of the increasing diversity of
the lower taxa; the general pattern is not bottom to top. Rather, the dominant
pattern is top to bottom, contrary to theory. As paleontologists Douglas
Erwin, James Valentine, and John Sepkoski describe the situation:
The fossil record suggests that the major pulse of diversification of
phyla occurs before that of classes, classes before that of orders, and
orders before families. This is not to say that each higher taxon originated
before species (each phylum, class, or order contained at least one species,
genus, family, etc. upon appearance), but the higher taxa do not seem to
have diverged through an accumulation of lower taxa (Erwin, Valentine, and
Sepkoski, 1988).

FIGURE 4: Pervasive pattern of natural history: Disparity precedes
diversity.
"We may acknowledge a central and surprising fact of life's history
- marked decrease in disparity followed by an outstanding increase in diversity
within the few surviving designs."
Stephen Jay Gould, Wonderful Life, 1989, p. 49
In another article Valentine and Erwin review hypotheses as to the
mode of origin of animal body plans for consistency with the fossil evidence.
They conclude that both Darwinian gradualism and punctuated equilibrium
are inadequate to account for the appearance of invertebrate body plans
and their major modifications:
The models we consider are of three sorts: those that extrapolate processes
of speciation to account for higher taxa via divergence, those that invoke
selection among species, and those that emphasize that many higher taxa
originated as novel lineages in their own right, not only as a consequence
of species-level processes. It is in this latter class of model that we
believe the record favors (Valentine and Erwin, 1985, p. 71).
If large populations have gradually evolved there should be unmistakable
evidence in the fossil record, yet it is simply not found.
... many of the large populations should have been preserved, yet we
simply do not find them. Small populations are called for, then, but there
are difficulties here also. The populations must remain small (and undetected)
and evolve steadily and consistently toward the body plan that comprises
the basis of a new phylum (or class). This is asking a lot. Deleterious
mutations would tend to accumulate in small populations to form genetic
loads that selection might not be able to handle. Stable intermediate adaptive
modes cannot be invoked as a regular feature, since we are then again faced
with the problem of just where their remains are. We might imagine vast
arrays of such small populations fanning continually and incessantly into
adaptive space. Vast arrays should have produced at least some fossil remains
also. Perhaps an even greater difficulty is the requirement that these arrays
of lineages change along a rather straight and true course --- morphological
side trips or detours of any frequency should lengthen the time of origin
of higher taxa beyond what appears to be available. Why should an opportunistic,
tinkering process set on such a course and hold it for so long successfully
among so many lineages?
We conclude that the extrapolation of microevolutionary rates to explain
the origin of new body plans is possible, but does not accord with the primary
evidence (Valentine and Erwin, 1985, pp. 95, 96).
The model of punctuated equilibrium or species selection attempts to account
for the lack of evidence by relying primarily on the evolution of small
isolated populations which would have a diminished chance of leaving a fossil
record. This scenario has its difficulties, however, as Valentine and Erwin
point out:
The required rapidity of the change implies either a few large steps
or many and exceedingly rapid smaller ones. Large steps are tantamount to
saltations and raise the problems of fitness barriers; small steps must
be numerous and entail the problems discussed under microevolution. The
periods of stasis raise the possibility that the lineage would enter the
fossil record, and we reiterate that we can identify none of the postulated
intermediate forms. Finally, the large numbers of species that must be generated
so as to form a pool from which the successful lineage is selected are nowhere
to be found. We conclude that the probability that species selection is
a general solution to the origin of higher taxa is not great, and that neither
of the contending theories of evolutionary change at the species level,
phyletic gradualism or punctuated equilibrium, seem applicable to the origin
of new body plans (Valentine and Erwin, 1985, p. 96).
This evidence further substantiates the proposition that minor lower-level
evolutionary change cannot be extrapolated to account for major evolutionary
change. This appears to be true for the both the tortoise and the hare,
Darwinian gradualism and punctuated equilibrium. The tortoise is far too
slow to account for the fossil evidence and the hare spends far too much
time in stasis.
Darwin admitted that the geological evidence was the "most obvious
and gravest objection which can be urged against my theory". At the
time, he was primarily concerned with the lack of transitional forms in
the fossil record. Today, those concerns are compounded by fewer transitional
forms than Darwin had in his day, and by the systematically upside-down
order of geological succession. Darwin was ironically prophetic in stating
that the facts can lead to conclusions directly opposite to those at which
he arrived.
Roger Lewin further describes the origin of most major body plans in the
Science Research News report, "A Lopsided Look at Evolution":
Described recently as "the most important evolutionary event during
the entire history of the Metazoa," the Cambrian explosion established
virtually all the major animal body forms -- Bauplane or phyla -- that would
exist thereafter, including many that were 'weeded out' and became extinct.
Compared with the 30 or so extant phyla, some people estimate that the Cambrian
explosion may have generated as many as 100. The evolutionary innovation
of the Precambrian/Cambrian boundary had clearly been extremely broad: "unprecedented
and unsurpassed," as James Valentine of the University of California,
Santa Barbara, recently put it (Lewin, 1988).
Lewin then asked the all important question: "Why, in subsequent
periods of great evolutionary activity when countless species, genera, and
families arose, have there been no new animal body plans produced, no new
phyla?" (Lewin, 1988).
If neo-Darwinian theory is true, why should
the Cambrian contain a greater number of body plans than exist today, particularly
with such low species diversity? Figures 5 and 6 graphically illustrate
the situation.
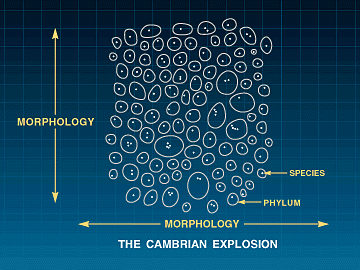
FIGURE 5: The Cambrian Explosion
The sudden appearance of between 50 and 100 disparate body plans with extremely
low species diversity supports the conclusion that neither gradual Darwinian
evolution nor lower taxon-level punctuations can account for the origin
of the higher taxa and the major body plans. In the history of life on earth,
disparity typically precedes diversity.

FIGURE 6: The Present
"We may acknowledge a central and surprising fact of life's history
- marked decrease in disparity followed by an outstanding increase in diversity
within the few surviving designs." (Stephen Jay Gould, 1989)
Lewin's question leads us to an even more important question. What mechanisms
have prevented major evolutionary change from occurring over the past 500
million years? Why did the origin of the phyla appear to have stopped first,
followed by classes and then orders?
One rather convincing explanation that has been offered for the pattern
is based upon the fact that any major novelty that arises relatively quickly
through non-selective processes is likely to be poorly adapted. The more
novel the body plan is, the more susceptible it will be to elimination by
competition and therefore the more adaptive space will be required for it
to become established. In short, competition tends to inhibit the establishment
of higher taxa. Natural selection tends to inhibit major evolutionary change.